
Chem/BMSE 259
Molecular Biology and Protein Purification Graduate Laboratory

|
Chem/BMSE 259
|
|
Prof. Rick Dahlquist
Office: Chem 1126C, Phone: 893-5326
E-mail: dahlquist@chem.ucsb.edu, Website: http://www.chem.ucsb.edu/people/faculty/dahlquist/index.shtml
Prof. Deborah Fygenson
Office: Broida 2409, Phone: 893-2449
E-mail: deborah@physics.ucsb.edu, Website: http://www.physics.ucsb.edu/~deborah
Dr. Kalju Kahn
Office: PSB-N 1511, Phone: 893-6157
E-mail: kalju@chem.ucsb.edu, Website: http://www.chem.ucsb.edu/~kalju
Andrew Bonham
Office: Chem 1148, Phone: 893-8283
E-mail: bonham@gmail.com
The UCSB summer biochemistry lab course is designed to introduce participants whose primary field is not biochemistry to modern techniques of molecular cloning, mutagenesis, and protein purification. Course participants will carry out a project similar to that performed in many contemporary biochemistry and molecular biology laboratories. In the first part of the project, participants clone a gene for a protein involved in bacterial chemotaxis into a modern expression vector and change specific amino acids in this protein by site-directed mutagenesis of the corresponding plasmid DNA. In the second part of the project, participants express the wild-type and mutated proteins in the bacterial cells and purify the overexpressed proteins by means of column chromatography. In the third part of the project, participants characterize the wild-type and mutant proteins by electrospray mass spectrometry and perform binding studies using fluorescence and nuclear magnetic resonance spectrometry.
The theory manuals and chapters of the operations manual can be downloaded here in the PDF format. Please note that you can follow hyperlinks that are in the PDF files by clicking on the link.
| Experiments | Download Adobe Acrobat Here | Acrobat |
| General | Theory Manual, Summer 2006 | |
| General | Mass Spectrometry: Background | |
| Practice | Agarose Gel Electrophoresis of DNA Isoforms | |
| Cloning | Operations Manual: Subcloning CheY from pCW into pET-28b | |
| Purification | Operations Manual: Purification of CheY Protein |
Many experiments in this course involve use of materials or equipment for which manufacturers have provided nicely illustrated operations manuals. Normally, you want to obtain the most up-to-date versions of these manuals from the manufacturer's website. Copies of some of these documents are provided here for your convenience.
| Literature | Technical Literature in PDF | Acrobat |
| Cloning | Stratagene StrataPrep DNA Gel Extraction Kit: Manual | |
| Cloning | Novagen pET System Manual | |
| Cloning | Novagen pET-28 Map | |
| Cloning | Stratagene XL1 Competent Cells: Manual | |
| Cloning | Stratagene XL10 Ultracompetent Cells: Manual | |
| DNA Purif. | QIAprep Miniprep Handbook | |
| Mutagenesis | Stratagene QuikChange© Kit: Manual | |
| Protein Purif. | BioRad Bradford Assay Manual | |
| Protein Purif. | BioRad Chromatography Guide | |
| Protein Purif. | BioRad EconoPump Manual | |
| Protein Purif. | BioRad Fraction Collector Manual | |
| Protein Purif. | BioRad MiniProtean 3 Manual | |
| Protein Purif. | Stirred Ultrafiltration Cell Manual | |
| Protein Purif. | Ultrafiltration Disks Manual |
| Lectures | Download Adobe Acrobat Here | Acrobat |
| Cloning | Structure and Function of DNA | |
| Cloning | DNA Technologies | |
| Cloning | Display Technologies (Dr. Daugherty) | No PDF |
| Characterization | Biological Mass Spectrometry (Dr. Pavlovitch) | |
| Characterization | Biological NMR Spectrometry (Dr. Kahn) |
| Publications | Suggested Scientific Publications in PDF | Acrobat |
| Engineering | "From genes to machines: DNA nanomechanical devices" by Nadrian Seeman | |
| Characterization | "Electrospray: Principles and Practice" by Simon Gaskell | |
| Characterization | "Structure Determination of Biological Macromolecules in Solution Using NMR Spectroscopy" by Gerhard Wider |
Participants in the UCSB Summer Biochemistry Lab Course clone, mutate, purify, and characterize mutant forms of the CheY protein. This protein is an important regulator of bacterial chemotaxis. During the first week, participants sub-cloned the cheY gene from the pCW plasmid into the pET-28b vector in order to create a His-tagged recombinant protein. During the second week, the recombinant protein was purified using Ni-affinity chromatography. During the third week, mutant forms of CheY were created using PCR-based site-directed mutagenesis technology and the expressed proteins were characterized by mass spectrometry. The binding of FliM peptide to CheY was characterized by fluorescence spectrophotometry in order to determine the ionic strength dependence of the binding constant.
| Description | Image |
|---|---|
PCR Cloning of CheY One of the first steps in subcloning the cheY gene involves PCR-amplification of the gene out from the parent pCW plasmid. Two primers, complementary to the 5' and 3' ends of the cheY gene were designed such that the primers introduced a Nco I restriction site at the N-terminus of the protein and eliminated a stop codon at the C-terminus. The PCR product was gel-purified and digested with Nco I and Xho I restriction endonucleases to obtain a fragment suitable for ligation into pET-28 vector. The image on the right shows purified digested PCR products from participants in the course. |
 |
Ligation of CheY with pET-28b The purified fragment of DNA containing the cheY gene was ligated to pET-28 vector that was linearized with NcoI and XhoI restriction endonucleases and gel-purified. Ligation reaction contained about 20 ng of CheY fragment and 80 ng of linearized pET vector. The image on the right shows the DNA composition of the ligation reaction after incubation with T4 DNA ligase. Successful ligation in some samples is evident by the appearance of slower-migrating circular DNA (e.g. lanes 5, 6, and 14) Due to the small quantity of DNA analyzed, it is possible that successful ligation products were also present in other samples. |
 |
Bacterial Transformation Competent E. coli (Stratagene XL10-Gold strain) were transformed with the ligation product using the heat-shock method. Transformed bacteria were plated on kanamycin-containing agar plates to select for clones that incorporated the ligation product. The image on the right shows one such agar plate after 16-hour incubation at 37 degrees. Other plates can be seen in the image gallery. Note that not all the colonies have incorporated the cheY gene as infrequent self-ligation of the pET-28 also produces antibiotic-resistant bacteria. |
 |
DNA Analysis Plasmid DNA from several colonies was purified and analyzed. Purified plasmid was digested with Nco I and Xho I restriction endonucleases to verify the presence of the cheY gene. Double digestion of the recombinant plasmid will yield a small fragment of about 400 bp and a large fragment of approximately 5.1 kbases. The image on the right shows results from such double digestion. The first lane contains 1 kb marker ladder, the second lane undigested purified plasmid, and the last lane reference cheY gene. Other lanes contain about 180 ng of digested DNA. Unfortunately, because of the small size of the cheY-containing fragment, the total amount of DNA in this fragment is too small for a reliable detection on the ethidium bromide-stained gels. |
 |
DNA Analysis However, the large fragment that consists of the rest of the pET-28 could be easily distinguished from the linearized recombinant DNA when smaller sample loads are used. The image on the right shows such restriction analysis of a recombinant DNA sample (sample from lane 14 on the previous image). Here, lane 1 contains 1 kb marker ladder, lane 4 undigested plasmid, lane 5 a plasmid treated with the Nde I restriction enzyme (no cuts expected in recombinant DNA), the sample in lane 6 was digested with Nco I (one site expected), the sample in lane 7 was digested with Xho I (one cut expected), and the sample in lane 8 was double digested with Nco I and Xho I (two cuts expected). Lanes 10-14 have unrelated samples. |
 |
Protein Expression Analysis Purified DNA from XL10-Gold cells was used to transform BL21(DE3) cells. A single colony was picked from each plate and the expression of the CheY protein was induced by adding 0.4 mM IPTG to the small liquid culture. The protein expression was checked by analyzing the protein composition using SDS polyacrylamide page electrophoresis on precast horizontal minigels. The recombinant His-tagged CheY migrates slightly faster than lysozyme, which was used for cell lysis. Thus, a double-band in the 15 kDa region indicates a successful expression of C-terminally His-tagged CheY. Lanes 1 and 8 have reference His-tagged CheY, lanes 3, 5, and 7 show expression of C-terminally tagged CheY while sample in lane 6 may have CheY without the C-terminal His-tag. |
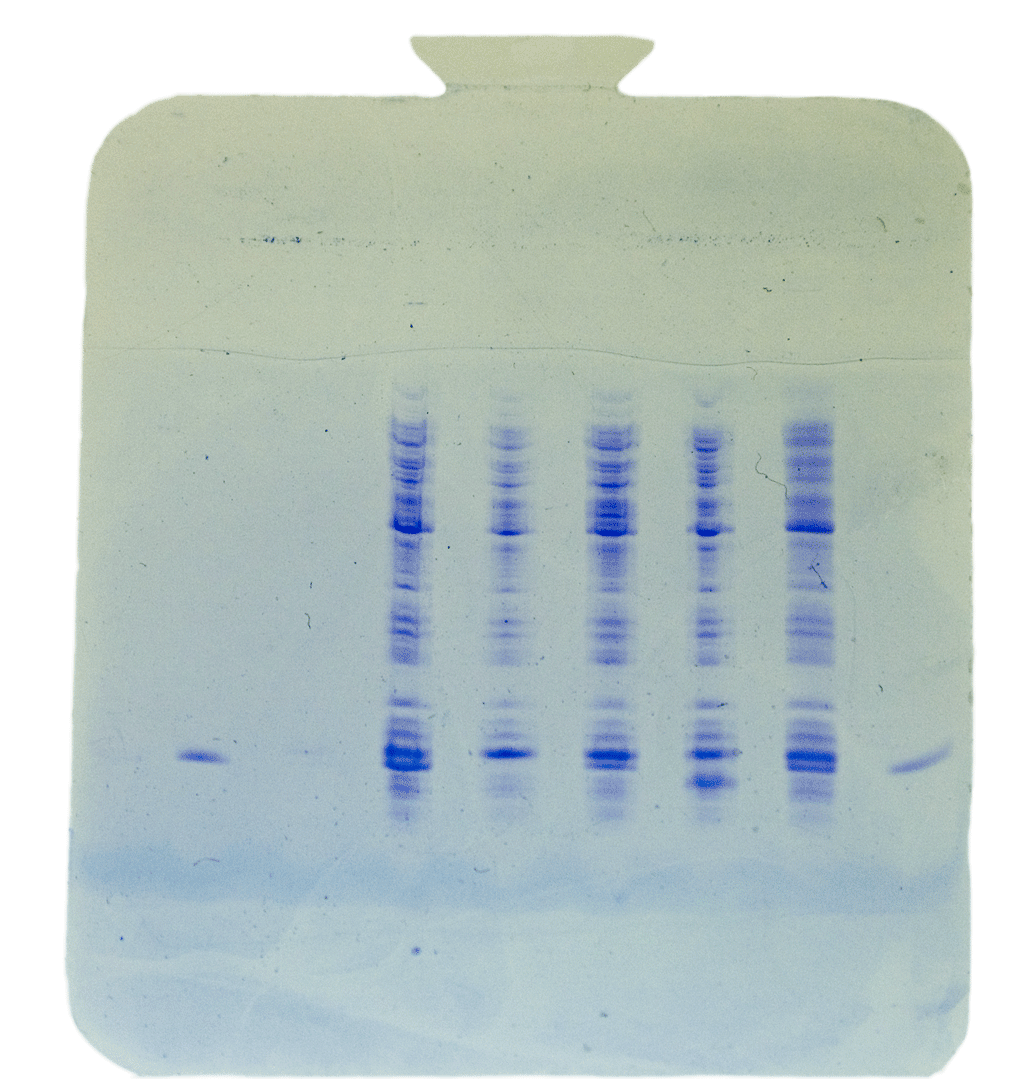 |
Protein Purification Cells expressing CheY protein were grown in large liquid cultures and lysed. The cell lysate was fractionated by adding ammonium sulfate to 65% saturation. The protein pellet was redissolved in phosphate buffer and dialyzed against 50 mM sodium phosphate, 300 mM NaCl, 10 mM imidazole, pH 8. The His-tagged CheY protein was bound to nickel-NTA agarose resin and eluted with a gradient of imidazole. The purity of eluted fractions was determined by vertical SDS-PAGE. The image on the right shows one Coomassie-stained SDS gel in which the second lane contains crude cell extract and the fourth lane contains purified CheY. The last lane has molecular weight markers. Another gel showing purified CheY in several fractions can be seen here. |
 |
Protein Immunodetection The presence of CheY in fractions was assayed by a sensitive immunodetection method in which fractions blotted to nitrocellulose membrane were probed with the Ni-NTA conjugated to horseradish peroxidase (HRP). The Ni-NTA binds to His-tagged proteins and HRP catalyzes the formation of an insoluble purple dye at these locations. Thus, samples containing His-tagged proteins will yield a purple dot on the paper. The image on the right shows one such immunoblot indicating that CheY eluted in fractions 24-27. Another immunoblot can be seen here. |
 |
Site-Directed Mutagenesis A double mutant of CheY was created from the K13D/I95V/Y106W triple mutant by changing the residue 95 back to isoleucine using site-directed mutagenesis Eighteen rounds of PCR amplification with annealing at 53-55 degrees were performed and the PCR product was analyzed on the agarose gel after Dpn I treatment. The image on the right shows one such gel. The first two lanes contain 1 kb size markers, the third and sixth lane have CheY double mutants, the fifth lane has control PCR product, and the seventh lane contains a sample from an unsuccessful PCR reaction. |
 |
Transformation with Mutant DNA The competent E. coli cells (Stratagene BL21(DE3) strain was transformed with the product of the site-directed mutagenesis reaction. Transformed bacteria were plated on kanamycin-containing agar plates to select for clones that took up the recombinant. The image on the right shows one such agar plate after 16-hour incubation at 37 degrees. Note that the presence of colonies in this experiment merely indicates that the bacteria had acquired resistance for kanamycin; the clones may or may not express His-tagged CheY. |
 |
Colony Expression Analysis The expression of His-tagged CheY by each colony was assayed by blotting the colonies onto nitrocellulose paper and inducing protein expression by growing blotted colonies on agar plates containing 0.25 mM IPTG. The membrane-bound cells were lysed and the presence of CheY was detected with Ni-NTA-HRP conjugate. The Ni-NTA binds to His-tagged proteins and HRP catalyzes the formation of an insoluble purple dye at these locations. Thus, cells that expressed His-tagged CheY yield a purple dot in the location of the colony. The image on the right shows the nitrocellulose paper with cells taken from the same agar plate that is shown on a previous image. |
 |
Purification of Mutant CheY A colony expressing double mutant CheY was picked from the plate above and used to grow 600 mL liquid culture, Protein expression was induced by adding 0.4 mM IPTG in the late phase of the growth. Bacteria were lysed using French press and the His-tagged mutant CheY was purified with Ni-NTA spin columns. The image on the right shows SDS-PAGE minigel of conventionally purified CheY in the first lane, five samples of the spin-column-purified CheY, the flow-through, and the cell extract before purification. Note that that while spin-columns provide a rapid and convenient purification, noticeable amount of impurities elute with CheY after two washes. |
 |
Mass Spectrometry of CheY The identity of the triple mutant CheY was analyzed by electrospray time-of-flight mass spectrometry. Purified protein was dialyzed extensively against deionized water and 20 µmolar sample was mixed with an equal volume of acetonitrile to facilitate evaporation of the solvent. The protein was ionized by addition of formic acid to a final concetration of 0.5 %. The image on the right shows the observed mass spectrum with a characteristic charge envelope arising from positively charged ions of intact CheY. Each major peak differs from the previous one by one charge unit. Analysis of this spectrum yields molecular mass of 15053 for the neutral protein molecule. This value is in a good agreement with the expected value for the triple mutant of CheY. |
 |
Binding of FliM: Fluorescence Study The biological function of CheY in bacterial chemotaxis is due to its ability to temporarily interact with the motor protein FliM. Participants in the course characterized the binding of the FliM peptide to the purified CheY protein using fluorescence spectroscopy. The binding of FliM will lead to significant quenching of the intrinsic fluorescence of CheY because the environment around the emitting tryptophan residue changes significantly upon binding. The extent of fluorescence quenching is proportional to the fraction of bound CheY. Analysis of fluorescence quenching as function of FliM concentration allows determination of the dissociation constant for the CheY-FliM complex. The image on the right shows results from one such experiment, yielding Kd value of about 3.0 µM. |
 |
NMR Characterization of CheY The purified triple mutant of CheY was characterized by nuclear magnetic resonance spectrometry. NMR is a valuable tool for determination of structures of biological macromolecules. One of the steps in determination of protein structures by NMR is assignment of observed resonances based on various multidimensional NMR experiments. The image on the right shows an aliphatic region of the heteronuclear single quantum coherence (HSQC) spectrum showing the 1H-13C correlations. The methyl group of residue Val 95 that is involved in the binding of FliM can be identified as a signal that in the free CheY appears at about 0.08 ppm. |
 |
NMR Characterization of FliM binding The binding of FliM to the purified triple mutant of CheY was studied by nuclear magnetic resonance spectrometry. NMR allows to monitor which CheY residues are involved in binding by comparing chemical shifts of protons in the free and bound forms of the protein. The image on the right shows an aliphatic region of the heteronuclear single quantum coherence (HSQC) spectrum showing the 1H-13C correlations. The methyl group of residue Val 95 that is involved in the binding of FliM appears at about 0.08 ppm in the 1H NMR spectrum in the absence of FliM peptide but shifts to the -0.44 ppm in the FliM bound form. |
 |
NMR Lineshape Analysis The observed NMR spectrum of CheY-FliM complex suggested that the dissociation rate of the complex is slow in the NMR chemical shift time scale. Specifically, the appearance of a broad (40-60 Hz) peak at -0.44 ppm and disappearance of a sharp (ca 18 Hz) peak at +0.08 ppm in the 600 MHz NMR instrument suggests that the half-life of the complex is well over one millisecond. Detailed simulation of the NMR line shapes based on the McConnell equations provided an estimate for the dissociation rate constant in the range of 30 to 70 s-1. |
 |
Crystallization of CheY Crystallization trials were set up to see if the triple mutant CheY could be crystallized. Protein crystallization is a required step for the determination of high-resolution structures by X-ray crystallography. Solutions of purified CheY were mixed with different precipitating agents in the hanging drop method. After two days, drops were examined under the polarizing microscope in search of protein crystals. Similar experiment was performed with lysozyme, which, as shown on the image on the right, readily crystallized. CheY, however, tended to precipitate out of the solution as amorphous or polycrystalline material not suitable for structure determination. Possibly, two small protein crystals were observed under two different conditions (CheY crystal 1; CheY crystal 2). |
 |
Pymol Examples here at UCSB *with scripts*
Electronic Journals at UCSB
Google Scholar Web Search Engine
Biology Workbench
ExPASy Proteomics Server
Protein Data Bank
Periodic Table of the Elements
Cell and Molecular Biology Protocols Online
UCSB General Catalog
UCSB Campus Map
UCSB Gold Login
UCSB Umail Access
UCSB Environmental Health and Safety
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}